Favoritos de dgborin
Favoritos de dgborin

Fotos / Sonidos

Observ.
ian_gardinerDescripción
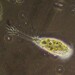
Habitat temporary grassland pool.
Image 1 - Dorsal view.
Image 2 - Ventral view.
Image 3 - Caudal lamina.
Image 4 - Eyes.
Image 5 - Nuchal organ (between the eyes).

Fotos / Sonidos

Observ.
peptolabDescripción
Paramecium nephridiatum V. Gelei, 1925 from the brackish estuarine pond Pussy's Pond, an offshoot of Acabonac Harbor. The population arose after feeding the culture with boiled wheat seeds. Imaged in Nomarski DIC on Olympus BH2S using SPlanapo 40 0.95 and SPlan 100 1.25 oil objectives plus variable phone camera cropping on Samsung Galaxy S9+.
The cells measure from 100-165 um in length. There is a large oval macronucleus usually situated slightly anterior to the cell equator and 3-4 small endosomal-type micronuclei can be observed with exact focus in the anterior end of the cell remote from the macronucleus- an important specific character of P. nephridiatum according to Fokin et al 1999 (1) The oral apparatus is situated posterior to the macronucleus with the cytostome roughly equatorial. There is an inconspicuous tuft of long caudal cilia. The two contractile vacuoles, one anterior and one posterior, have up to 15 short radiating canals and 2-3 excretory pores- another important specific characteristic of P. nephridiatum (1). There is a dense investment of subpellicuar extrusomes. The pellicle is divided into rectangular spaces on optical sectioning of the surface, a feature not mentioned by Fokin et al 1999. These features point strongly to P. nephridiatum. However, the more posterior location of the oral opening (as opposed to slightly anterior to the middle of the cell) and the peculiar pellicular division into rectangular spaces suggest that this might be a cryptic species of P. nephridiatum. Indeed, Przyboś et al 2019 describe 6 cryptic species within P. nephridiatum (2)
The cell shape with its rounded posterior end, and broad anterior part is of the "bursaria" type (Wichterman 1986) or "woodruffi" type according to Jankowski (1969). The oral opening is situated a little anterior to the middle of the cell. The cell surface is uniformly ciliated except for several long caudal cilia located close to the posterior end of the dorsal side, but not on the top. The abundant subpellicular trichocysts are distributed uniformly. Specimens were about 145 x 47 um long in vivo but shrank 10% after the silver nitrate impregnation. In silver-impregnated specimens, there were ~38 rows of ventral kineties and ~35 dorsal kineties. The preoral suture is distinct, but the postoral suture is very obscure. The cytoproct is situated in the posterior third. The vestibular zone is conspicuous and is terminated by the distinctive shape of the buccal opening. Two peniculi and open quadrulus are located on the dorsal and left wall of the buccal cavity. The endoral membrane is situated along the entire right edge of the buccal opening but its dikinetids are not recognizable in all specimens. It has 15 dikinetids on the average. The buccal cavity size various around 30 um. On the dorsal side of the body the contractile vacuole pores are very distinctive both on impregnated specimens as well as in living cells. Usually, both contractile vacuoles have more than one pore each, typically two or three. However, we have found two stocks (WCh-1 and WS-12) where one of the contractile vacuoles quite often (up to 50%) had only one pore. Both contractile vacuoles usually have 8-14 collecting canals, ten on average. Numerous crystals were very often found in the cytoplasm, but their quantity and location varied, probably depending on the culture conditions. The nuclear apparatus is located in the anterior part of the cell. One slightly ellipsoid or ovoid macronucleus, ~30x36 um in living cell and ~17x23 um in stained cells, on average, resides just anterior to the equator of the cell. In Feulgen-stained cells the macronucleus has a very intense colour. The three to four spherical micronuclei of the "endosomal" type (Fokin 1997), ~3 um in diam. (on average) are distributed irregularly along the anterior part of cell. Endocytobiotic bacteria are often found in the cytoplasm (Fokin 1989) and can also be found in the perinuclear space and in the macronucleus (Fokin 1989; Fokin and Sabaneyeva 1997)" (1).
"Swimming behavior - During swimming this species spirals on its long axis in both directions (Fokin 1987). We could not find any simple triggers (food, time) for changing this swimming direction. Typically, "left spiral swimmers" and "right spiral swimmers" were present at the same time in the culture. During the several years of investigation of this trait there was some preference of the cells from the same stocks to spiral in the left direction. Remarks- Gelei (1925) described a new species of Paramecium, Paramecium nephridiatum, based on the material which he had found in his laboratory aquarium. This population was in fact a mixture of the new species and Paramecium caudatum (Gelei 1938). This was the reason why some features of the new ciliate were similar to P. caudatum so that no one recognized this new species in nature, though reference to P. nephridiatum was made by Kahl (1931) and Kalmus (1931). Gelei (1938) redescribed the species from a native population (Tisza River, Szeged, Hungary) using a "clean culture". For unknown reasons, this new description did not attract the attention of protozoologists and in all subsequent reviews, P. nephridiatum was considered a nonvalid species (Vivier 1974; Wichterman 1953, 1986) even when the article of Gelei (1938) was listed in the references. Only once was P. nephridiatum mentioned in a short abstract as a species living in Florida, USA (Bovee 1983), although characters of the species were not listed in this publication. Since 1983, one of us (S. F) has repeatedly collected a species of Paramecium with multiple contactile vacuole pores, which is a distinctive trait of P. nephridiatum, although it was considered for a time as a feature of Paramecium woodruffi (Agamaliev 1983; Fokin 1986), Jankowski (pers. commun.)" (1).
"Occurrence and ecology- A number of stocks of P. nephridiatum were isolated from the sea shores of northern Europe: the North, Baltic, White, and Barents Sea coasts. It was detected during sampling in Woods Hole, MA, USA, Atlantic Ocean and on Sakhalin Island, Sea of Okhotsk. The salinity of these samples varied from 1.5-32 % The species was also found in a fresh-water body in Jerusalem Zoo, Israel. Samples were taken mainly during the summer, from mid-April (Wood Hole) to November (North Sea coast).
Sampling of the same wild population of P. nephridiatum (Sredny Island, White Sea, Russia) has been repeated every year since 1990 to observe long-time changes in the population, as well as the euryhaline ability of the species. This population was present at salinities from 4-35% and in the temperature range from 10-25” C. Very often the populations of P. nephridiatum occurred at the lower limit of oxygen concentrations. They were mainly feeding on bacteria. In the same samples these other ciliates were usually found: Prorodon sp., Frontonia marina, Metopus sp., P. calkinsi, P. woodruffi, and sometimes P. duboscqui" (1).
- Sergei I. Fokin, Thorsten Stoeck, and Helmut J. Schmidt; Rediscovery of Paramecium nephridiatum Gelei, 1925 and its Characteristics. J.Eukaryot.Microbiol. 46(4):416-426, 1999
- Evaluation of the molecular variability and characteristics of Paramecium polycaryum and Paramecium nephridiatum, within subgenus Cypriostomum (Ciliophora, Protista). Ewa Przyboś, Maria Rautian, Alexandra Beliavskaia, Sebastian Tarcz . Mol Phylogenet Evol . 2019 Mar:132:296-306
Fotos / Sonidos
Qué
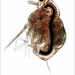
Pulga de Agua (Moina macrocopa)Observ.
ian_gardinerDescripción
Image 1 - Female.
Image 2 - Bifid distal tooth on the postabdomen, as with all North American Moina with the exception of M. hutchinsoni (single tooth).
Image 3 - A series of teeth (arrowed) on the outer surface of the anterior seta of the penultimate segment of the first leg - a feature unique to this species.
Fotos / Sonidos

Observ.
closterium_mysteriumDescripción
Video: https://youtu.be/5APrHApGLXc
Sampling location: A tardigrade was collected from a sample of Trentepohlia umbrina found on a tree bark within the city limits.
Date and time of collection: 27 Aug 2023 at 10 AM
Date and time of observation: 27 Aug 2023 at 3 PM
The sample was stored at room temperature in a plastic container.
T. umbrina observation: https://www.inaturalist.org/observations/198383520

Fotos / Sonidos

Observ.
mnold1Descripción
Mag. 400x
- A water sample taken on 03/31/2024, from a small vernal stream in a wooded area, using a small sample bottled attached to an extension pole. Air temp 46F.
I'm not certain about the ID, but will check with geraldojpr and roman_romanov. Most fascinating about this specimen is that one semi-cell is 3-radiate (3 lobes) and the other is 6-radiate. An intermediate polymorph! Will its progeny be 3- or 6 radiate or a hybrid of both? Interesting! S. margaritaceum is noted to be "variable in the in the number of radial processes, , with 4-6 being the most often found" (https://www.outerhebridesalgae.uk/desmids/desmid-species.php?id=359) ... I could not find a reference noting 3-radiate S. margaritaceum. The first composite image shows the specimen in lateral view, then beginning to roll backwards. The second composite of 9 photos shows the same specimen making a ~180⁰ rotation about a slanted axis. (Panels 8 and 9 vary only in depth of field, not position of the specimen.) Too cool!
For more reference images of S. margaritaceum see:
Fotos / Sonidos

Observ.
mnold1Descripción
Mag. 400x
- A water sample taken on 03/31/2024, from a small vernal stream in a wooded area, using a small sample bottled attached to an extension pole. Air temp 46F.
3 Cosmarium of ~equal size and shape. The diagnosis was made based on images here http://www.digicodes.info/Cosmarium_punctulatum_var_subpunctulatum.html and here https://www.desmids.nl/maand/english/cosmarium_punctulatum.html. The reference images mimic the variation in my 3 specimens, especially regarding the variable flatness of the apices. The apical view of the second specimen is very similar to an image shown at the first link and also here https://www.algaebase.org/search/images/detail/?img_id=19326.
Fotos / Sonidos

Qué
Pulga de Agua (Daphnia pulex)Observ.
ian_gardinerDescripción
Male.
Habitat: temporarily flooded margin of small, permanent pond.
Fotos / Sonidos
Qué
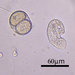
Género ColpodaObserv.
crseaquistDescripción
Gathered dry leaves on 2024-02-23 and stored in water.
Two daughters emerging from a division cyst:
- Initial observation of cyst with another individual
- 2 minutes later
- 18 minutes later
- 77 minutes later
- 84 minutes later
- 140 minutes later
- 153 minutes later
- 154 minutes later 1st daughter emerges
- 155 minutes later
- 162 minutes later
- 163 minutes later 2nd daughter emerges
- 2nd daughter swims away
Video: https://youtu.be/vntikKDmSDE

Fotos / Sonidos
Observ.
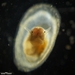
closterium_mysteriumDescripción
Video: https://youtu.be/Pmvk5Baer_U
Sampling location: A water sample was collected from Istok Lake near the shore, where snags and aquatic vegetation were present.
Date and time of collection: 16 Jul 2023 at 3 PM
Date and time of observation: 20 Jul 2023 at 6 PM
The sample was stored at room temperature in a plastic container.
Fotos / Sonidos

Observ.
ian_gardinerDescripción
Habitat hypersaline lake.
Image 1 - Male, dorsal view.
Image 2 - Female, dorsal view.
Image 3 - Male head, frontal view, showing the distal segments of the second antennae to be i) flat, ii) broadly triangular, iii) with pointed apices directed medially.
Image 4 - Habitat.
The major solution salt in this lake is sodium carbonate (Cummings, J.M. 1940. Saline and Hydromagnesite Deposits of British Columbia, Bulletin No. 4. BC Dept of Mines: 160pp)
Fotos / Sonidos

Qué
Alonella excisaObserv.
ian_gardinerDescripción
Female.
Image shows the striae within the carapace polygons, characteristic of this species.

Fotos / Sonidos
Qué
Adineta steineriObserv.
closterium_mysteriumDescripción
Observation of a single specimen.
After rehydrating, the rotifer crawled for only a short time before returning to its contracted state, having found nothing of interest.
The sample was obtained from moss collected in the forest and stored at room temperature for three days prior to observation.
Video: https://youtu.be/38P-ilxFzfo
Fotos / Sonidos

Observ.
ian_gardinerDescripción
Habitat small, shallow lake.
Image 1 - Male, ventral view.
Image 2 - Female, ventral view.
Image 3 - Female, left lateral view, with eggs and two attached spermatophores.
Image 4 - Male 5th legs, posterior view, showing hyaline process on distal portion of inner margin of right exopod 1.
Image 5 - Male right 1st antenna, showing process on antepenultimate segment.
Image 6 - Female dorsal posterior view, showing 1) metasomal wings essentially symmetric, 2) the left lobe of the genital segment significantly smaller than the right, and 3) the lobes of the genital segment within the middle third of the segment - all points of difference between this species and the closely-related L. siciloides.


Fotos / Sonidos

Observ.
douchDescripción
Bright-field microscopy. Between barbs, on internal surface of detached Threskiornis molucca molucca primary feather.
Image 1: body
Image 2: anterior; upper focal-plane
Image 3: anterior; central focal-plane
Image 4: anterior; lower focal-plane
Image 5: posterior; upper focal-plane
Image 6: posterior; lower focal-plane
Image 7: mites on feather
Image 8: feather
Fotos / Sonidos

Observ.
ian_gardinerDescripción
Habitat small, permanent pond.
Image 1 - male, ventral view.
Image 2 - female, ventral view.
Image 3 - male 5th pair of legs, posterior view.
Image 4 - male right first antenna, showing apex with a pointed outwardly-produced process (hence denticornis = toothed horn).

Fotos / Sonidos

Observ.
ian_gardinerDescripción
Habitat small, temporary, grassland pool.
Image 1 - female, ventral view with eggs and two attached spermatophores.
Image 2 - male, ventral view.
Image 3 - male 5th pair of legs, posterior view, showing i) inner and distal processes on left exopod 2 forming a pincer-like structure, ii) spiniform process on outer distal corner of right basis.
Image 4 - male right first antenna, showing short, curved, spinous process at distal end of antepenultimate segment.
Image 5 - female posterior dorsum showing elongate outer sensilla on metasomal wings.
Image 6 - female 5th pair of legs.


Fotos / Sonidos

Observ.
closterium_mysteriumDescripción
Fig. 1-3: Valve view
Fig. 4-6: Girdle view
GIF and Fig. 7-9: A few images as the diatom rotates
A water sample was taken from the bank of the Vuoksi River. The sample was stored at room temperature and observed one day after collection.
Video: https://youtu.be/n7HxWvJmyp4
Observation of Entomoneis ornata https://www.inaturalist.org/observations/192091414
Fotos / Sonidos

Observ.
kenkDescripción
Specimen approximately 2mm. On the shell of a California Mussel (Mytilus californianus). The GIF is at 50% speed.
Fotos / Sonidos

Qué
Sida crystallinaObserv.
ian_gardinerDescripción
Male.
Habitat lake, marginal vegetation.
Fotos / Sonidos

Observ.
ian_gardinerFecha
Septiembre 28, 2011Descripción
Image 1 - male, ventral view.
Image 2 - female, ventral view.
Image 3 - male urosome, dorsal view.
Image 4 - male 5th legs, posterior view.
Image 5 - female 5th legs.
Fotos / Sonidos

Observ.
peptolabDescripción
An unusual flask-shaped contractile ciliate, an as yet undescribed haptorian possibly related to Chaenea, from the northernmost saprobic edge benthos of the spring-fed freshwater coastal pond at Ocean Dunes Apartments in the Atlantic Double Dunes Reserve. The sampling site is located 250 meters from the edge of the Atlantic Ocean. Imaged in Nomarski DIC on Olympus BH2S using SPlan 40x objective plus variable phone camera cropping on Samsung Galaxy S9+.
So far I have found two individuals, one of which is better for viewing the macronuclei but did not exhibit the contractile behavior. They resemble some sort of haptorid. Chaenea or Lacrymaria come to mind in several expert opinions. The inconspicuous mouth favors a Chaenea-like animal and the lack of a head all but rules out a lacrymariid though the spiral contractility and flask-shape resemble Lacrymaria. There is also a resemblance to the more narrow Litonotus or Amphileptus species such as Litonotus cygnus or Amphileptus procerus. However details of the neck region do not show pleurostomatid oral structure.
The larger of the two cells measures 84 um in length. There is a long narrow neck like region with two longitudinal ridges. The oral opening is not identified but very likely at the tip. The neck does not show a mane-like ciliation of the ventral neck but rather is uniform on both dorsal and ventral surfaces. The neck is held straight with no bend or curvature but is quite flexible. The plump posterior region has a tapered rounded posterior end containing a terminal contractile vacuole. There is no dome-shaped hump. The macronucleus is likely composed of multiple small scattered nodules (per Bill Bourland and Peter Vďačný opinions). This is better seen in the second smaller individual. The ellipsoid structure I initially interpreted as the macronucleus is likely a food vacuole. The ciliate swims forward in small arcs and often reverses direction swimming backwards for a similar distance similar to the motion of Lacrymaria as noted by Dr. Martin Kreutz, who thought it looks like a cross between Lacrymaria and Litonotus.
The most striking feature observed in this ciliate involves the contractile actions of the neck region. Normally, the neck is narrow tapering to a point anteriorly with two longitudinal ridges and what we assume to be an inapparent mouth opening at the pointed tip. Occasionally, the neck begins to broaden and the ridges become spiralized which then contracts the neck and forms a spatulate highly spiralized twisted structure at the anterior end of which there is a gaping cavity (pharynx?). This seems to be feeding behavior but at no time did I observe the ciliate ingesting any prey. Perhaps the ciliate is irritated by the coverglass. The second smaller individual did not exhibit this contractility.
I am grateful to several experts for taking the time to examine my images of this most unusual ciliate. None of them have ever seen anything like it and feel that this is likely something new and undescribed. I excerpt their kind opinions and impressions below:
Bill Bourland: "it looks like a very bizarre chaeniid litostomatean- I can't quite make out any dorsal brush or extrusomes (probably there but not quite visible). The preoral dome is very tiny and inconspicuous like some Chaenia species- the contractility is really impressive- there may be multiple small macronuclear nodules- the flask-shaped body (Lacymaria-like) is unlike most Chaenia species which are more narrowly ellipsoidal or fusiform. Very likely something new:) Thanks for sharing it!
Peter Vďačný: "Hi Jeff, I have not seen anything like that before. It looks like a mixture of a pleurostomatid and Lacrymaria. It somewhat resembles Litonotus cygnus or Amphileptus procerus. However, the contracted state resembles Lacrymaria. There is no head at the top of the neck. This excludes the possibility that this ciliate is a lacrymariid. It might be a Chaenea-like haptorid too. Some species are highly contractile. I am not sure, but it is not a lacrymariid for sure. The mouth of Chaenea are at the top and pretty indistinct. It is not a Chaenea, but it might be its relative. The type of contraction is very unusual. Only the neck is so highly contractile, as in Lacrymaria".
"I am not sure about the MA, maybe many small scattered MA nodules. There could be 100-200 MA nodules scattered throughout the cytoplasm. There are Chaenea species with hundreds of nodules. When there are many small scattered nodules, it is extremely difficult to recognize them. MA nodules could be very small. They may appear as lipid droplets or inclusions, sometimes almost impossible to distinguish even with DIC. Your MA appears to me as a food".
"Chaenea is a very deep branching lineage that stemmed right after helicoprorodontids. Lacrymariids belong to the rapid terminal radiation of haptorids. So, I know that helecoprorodontids branched off first, then Chaenea followed possibly by rhynchostomatians. The rest is unknown. But didiniids and spathiidids, both are related. So, helicoprorodontids and chaeneids are the deepest haptorian lineages. Both are contractile. About half of lacrymariids is contractile as well. Some pleurostomatids. Very few dileptids. The rest of haptorians is mostly not contractile, there are exceptions. However, the deepest lineages are highly contractile".
Fotos / Sonidos

Observ.
peptolabDescripción
A most unusual vermiform ciliate, probably a new species or even a new genus, found in a biofilm that formed on a 10 day old sample from sulfidic fine sand estuarine intertidal benthos of Acabonac Harbor at Landing Lane. So far I have found two of them measuring 225 um and 265 um in length trapped in the biofilm. The mouth opening with the prominent, large extrusomes looks quite like Chaenea. However, most Chaenea species have many macronuclei distributed in the cytoplasm while in this species there is only one large central macronucleus. Indeed, I often find Chaenea species in biofilms. DIC on Olympus BH2 using SPlan 40x objective plus variable phone camera cropping on Samsung Galaxy S9+ revealed the following features:
The body measures 225 um and 265 um in length. Body is vermiform with transverse annular ridges/paratenes running the full length of the body imparting an earthworm like appearance. The body is quite flexible and maybe a little contractile in life but they contracted markedly as they died. The mouth is apical and a simple slit. The head is not distinctly set off from the rest of the body by a neck region and has long dorsal brush cilia and a pharyngeal basket that resembles a peculiar subapical "window" through which a bundle of extrusomes can be seen. In addition, the first 5 segments comprising the head have regularly arranged rows of short rods probably nematodesmata or perhaps a second shorter type of extrusome. The pellicle is uniformly divided into a textured ?diamond-shaped pattern but I do not see a mucous layer. The macronucleus, situated just posterior to the mid-point of the cell, is large and ellipsoid with two micronuclei (or maybe nucleoli) one at either end. There are long rod-shaped extrusomes randomly all through the body. The posterior end has a terminal CV and a few various sized food vacuoles.
I sent it to Peter Vdancny who wrote: " I have had a look at your great discovery! However, I do not think that this is a haptorid. In my opinion, this could be a prostome morphologically somewhat similar (related) to Lagynus. Despite this, you might be correct that this is a completely new phylogenetic lineage. Somatic kineties are arranged in paratenes (circles), as typical of many prostomes. Moreover, many prostomes also have toxicysts (derived from trichocysts), a terminal contractile vacuole, and a dorsal brush =brosse. All these features are the results of the convergent evolution of haptorids and prostomes. And hence both groups could be sometimes mixed. Lagynus is anaerobic (though free-living) and its anterior body part is also annulated".
"Lagynusids are typically elongated, the cell being divided into the “head”, annular neck-like region, and “trunk”, which is an unusual feature in prostomateans (Foissner et al. 1995; Jiang et al. 2021b; Sola et al. 1990). When established, Lagynusidae included only one monotypic genus Lagynus Quennerstedt, 1867, with Lagynus elegans (Engelmann, 1862) Quennerstedt, 1867 as the type species. Later, two species, Lagynus cucumis (Penard, 1922) Foissner, 1987 and Lagynus binucleatus Jiang et al., 2021, were added (Foissner 1987; Jiang et al. 2021b; Sola et al. 1990). In terms of 18S rRNA gene sequence data, only that of L. binucleatus are available (Jiang et al. 2021b). The lack of further molecular data has hindered the progress of phylogenetic studies of the family Lagynusidae. In the study of Jiang et al 2023 (1) Ithe authors provided novel morphological and morphometric data as well as the frst 18S rRNA gene sequences of four microaerophilic species of Lagynusidae. They found that some species of lagynusids deviate distinctly from the type species or lack important characteristics (e.g., a conspicuous neck-like region and rod-shaped extrusomes). They revised the genus Lagynus and proposed the establishment of two new genera, Penardella and Apolagynus, based on complete morphological observations. They described two new species, P. marina and L. minutus, and made two new combinations, A. cucumis and A. binucleatus" (1).
I have found Lagynus elegans, the type species of the Laynudidae with its 5-7 annulated ridges involving the neck region, many times in my freshwater garden pond but the annulation of the entire length of this marine vermiform Lagynus-like ciliate and its textured pellicle are most unusual as is the large macronucleus with two large micronuclei. I sent images to Dr. Hu, the corresponding author of the Lagynusidae paper below. One of his students replied, again pointing to my initial impression of Chaenea: "I have checked the images you provided, but only based on the current information, I cannot identify this species accurately. There is no doubt that its oral is located at the anterior end of the body, so it may belong to the class Prostomatea or Haptorida (Class Litostomatea). The brosse (structure and location) is the key to distinguish these two groups, further tests (e.g., protargol staining) are needed for identification.The attachments I send are two papers about Cheanea (class Litostomatea, order Haptorida), and I think your species is similar to this genus, especially the obvious extrusomes near oral".
Based on the constellation of available features, I think this mysterious beauty has more affinities to the newer marine genera in the family Lagynusidae Sola 1990 such as Apolagynus cucumis and Penardella marina than to Chaenea with its annular constrictions, oral extrusomes single compact macronucleus.
Morphological and molecular examination of the ciliate family Lagynusidae (Protista, Ciliophora, Prostomatea) with descriptions of two new genera and two new species from China. Limin Jiang, Congcong Wang, Saleh A. Al‑Farraj, Hunter N. Hines and Xiaozhong Hu. Marine Life Science & Technology (2023) 5:178–195

Fotos / Sonidos
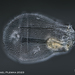
Observ.
plingfactoryDescripción
Asplanchna priodonta; spedieren with unknown parasites.
More on this here:
https://www.plingfactory.de/Science/Atlas/KennkartenTiere/Rotifers/01RotEng/source/Asplanchna%20priodonta.html

Fotos / Sonidos

Observ.
peptolabDescripción
Paramecium caudatum from the decomposing leaf debris in my long-neglected freshwater garden pond. Imaged in Nomarski DIC on Olympus BH2 using SPlan 20x and 40x objectives plus variable phone camera cropping on Samsung Galaxy S9+. The population ranges from 187-225 um in length. The cells are cigar-shaped with the posterior end bluntly pointed and with a group of slightly longer cilia compared to the otherwise uniform ciliation. Early microscopists likened its shape to that of a slipper, and commonly referred to it as the "slipper animalcule." The oral groove/ vestibulum is long and slightly oblique; buccal cavity with one endoral membrane and 2 peniculi. There is a single large ellipsoid macronucleus and 1 round compact micronucleus. " Using light microscopy, however, it is possible to discriminate only the Mi type of P. caudatum from the nucleus of other two species. P. caudatum always has the Mi with the so-called “cap”, a space without chromatin at the nuclear pole" (1). Two contractile vacuoles, each with radial canals, are present at the aboral surface which serve to excrete excess water taken up from the outside, regulating the water contents of the body. Radially distributed "collecting canals" give the contractile vacuoles a distinctive star-like shape. The cell is enclosed by a cellular envelope (cortex) densely studded with spindle-shaped extrusomes called trichocysts.
Discussion adapted from: https://en.wikipedia.org/wiki/Paramecium_caudatum
P. caudatum feed on bacteria and small eukaryotic cells, such as yeast and flagellate algae. the accumulated food particles, at the posterior end of the cytopharynx, are directed by the long cilia into the rounded, ball-like mass in the endoplasm called food vacuole. The food vacuoles are circulated by the streaming movement of the endoplasm which is called cyclosis. In hypotonic conditions (freshwater), the cell absorbs water by osmosis. It regulates osmotic pressure with the help of bladder-like contractile vacuoles, gathering internal water through its star-shaped radial canals and expelling the excess through the plasma membrane. When moving through the water, they follow a spiral path while rotating on the long axis.
Paramecium have two types of nuclei, a large macronucleus and a varying numbers of smaller micronuclei depending on the species. P. caudatum has 1 compact round macronucleus. The macronucleus controls somatic gene expression and the micronucleus controls reproduction. Like all ciliates, Paramecia reproduce asexually, by binary fission. During reproduction, the macronucleus splits by a type of amitosis, and the micronuclei undergo mitosis. The cell then divides transversally, and each new cell obtains a copy of the micronucleus and the macronucleus. Fission may occur as part of the normal vegetative cell cycle. Under certain conditions, it may be preceded by self-fertilization (autogamy), or it may follow conugation, a sexual phenomenon in which Paramecia of compatible mating types fuse temporarily and exchange genetic material. During conjugation, the micronuclei of each conjugant divide by meiosis and the haploid gametes pass from one cell to the other. The gametes of each organism then fuse to form diploid micronuclei. The old macronuclei are destroyed, and new ones are developed from the new micronuclei. Without the rejuvenating effects of autogamy or conjugation a Paramecium ages and dies. Only opposite mating types, or genetically compatible organisms, can unite in conjugation.
I observed a Paramecium in the early mid stages of transverse fission with formation of two oral anlagen and a beginning constriction of the cortex dividing into two cells.
Paramecium genus: biodiversity, some morphological features and the key to the main morphospecies
discrimination. Sergei I. Fokin. Protistology 6 (4), 227–235 (2010/11)
https://www.researchgate.net/publication/285129115


Fotos / Sonidos
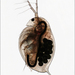
Qué
Daphnia similisObserv.
ian_gardinerDescripción
Female.
Habitat: saline lake.
Image 2 - Habitat.
Fotos / Sonidos

Observ.
ian_gardinerDescripción
Habitat temporary pool in roadside ditch.
Female showing distinctive rostrum.

Fotos / Sonidos

Observ.
peptolabDescripción
A gastrotrich from genus Turbanella from the superficial intertdal benthos of marine estuary Acabonac Harbor at Louse Point. The length is 375 um.
Turbanellidae is a family of worms belonging to the order Macrodasyida. Macrodasyida is an order of gastrotrichs.[1] Members of this order are somewhat worm-like in form, and not more than 1 to 1.5 mm in length. Macrodasyids are almost in entirely marine and live in the sediment in marine or brackish water, but two species have been discovered in freshwater. They can be distinguished from other gastrotrichs by the presence of two pores on either side of the pharynx, that allow excess water to be expelled during feeding. The body is dorsally flattened and there are tubular adhesive glands at both ends and on the lateral surfaces. These animals are detritivores and are hermaphrodites.
The genus Turbanella Schultze, 1853, contains 29 similar species. Morphological variability within and among species of Turbanella is extensive and has led to considerable synonymy (Hummon 2001). Several species of Turbanella are considered cosmopolitan (Wieser 1957; Ruppert 1977; Clausen 2000). "There are several species of Turbanella which give appearances of distributions on both sides of the Atlantic Ocean. They include: T. ambronensis Remane, 1943, T. bocqueti Kaplan, 1957 sensu Boaden, 1974, T. cornuta Remane, 1924, T. hyalina Schulze, 1857 and now T. amphiatlantica n. sp., but not, as we argue, T. mustela Wieser, 1957. Their locations are summarized in Hummon (2009), but see also d’Hondt (1971), Hummon (1997) and Todaro et al. (2008). All of these species are intertidal, sometimes shallow subtidal, and hence are expected to be euryosmotic over a wide range of salinities" (1).
- Turbanella amphiatlantica, a new species of Gastrotricha (Macrodasyida) from eastern North America and northwest Europe William D. Hummon and Jennifer L. Kelly. Meiofauna Marina, Vol. 18, pp. 61-70, Sept. 2011
Imaged in Nomarski DIC on Olympus BH2 using SPlan 20x objective plus camera phone cropping on Samsung Galaxy S9+.
Fotos / Sonidos

Observ.
ian_gardinerDescripción
Possibly the second record west of the Continental Divide, the first being http://n2t.net/ark:/65665/39d93e4e8-a728-44d6-b31e-86529dc0e56e
Habitat temporary pool with much vegetation.
Image 1 - male head showing fan-shaped antennae with three pointed tips.
Image 2 - male, ventral view.
Image 3 - male right second antenna, lateral view.
Image 4 - knob at base of distal segment of male left second antenna. Although Hartland-Rowe in his 1967 description of the species describes this knob as round, it's actually more-or-less pear shaped.
Image 5 - frontal view of male head.
Image 6 - habitat.
Fotos / Sonidos

Observ.
ian_gardinerDescripción
Female. Habitat small, shallow lake.
Image 2 - antennal setation 0-0-1-3 / 1-1-3, antennal spines 0-1-0-1 / 0-0-1, distal expansion of antennae with subapical ventral angulation.
Image 3 - dorsal serrations.
Image 4 - ventral-lateral view, showing the pronounced dorsal ridge.
Image 5 - postabdomen, showing that the dorsal rim is not bilobed (unlike some other members of this genus).
Image 6 - seta natatoria, showing the proximal segment to be longer than the distal, and to have no bristle-setae.
Image 7 - Habitat.

Fotos / Sonidos

Observ.
m_whitsonDescripción
I have my Bio 302L students at Northern Kentucky University to thank for this fun find. This amazing bacteria colony grew out of a water sample taken from the Lakeside Commons pond, directly behind the Campbell County Cooperative Extension offices. On 1/17, water and a little sediment was collected from the edge of the pond. On 1/19/2023 my microbiology students diluted a sample and spread plated it onto glycerol yeast-extract media, which is selective for gram-positives such as actinomycetes. This colony grew quietly on a 0.01 mL plate that had been forgotten in the fridge. Actinomycetes often have strange colony morphologies, so that's my guess on an ID for this. Neither I nor my colleagues have seen a colony like this before.

Fotos / Sonidos

Qué
Tortuga Pinta (Trachemys scripta ssp. elegans)Observ.
nelson_wisnikDescripción
in the absence of a trunk or stone to lean on, a capybara may be an option;
I have observed these two individuals do this twice;
see also
https://www.inaturalist.org/journal/nelson_wisnik/21258-the-friendly-capybara
Fotos / Sonidos

Observ.
ian_gardinerDescripción
Female. Jelly coat not visible.
Habitat: open water of lake.
H. glacialis is morphologically indistinguishable from H. gibberum, but their distributions are largely separate. The location of this specimen was very far from the area where both co-exist. See section 'Differential Diagnosis' on page 29 of the paper below:
https://www.researchgate.net/profile/Sarah-Adamowicz/publication/287753132_Three_New_Cryptic_Species_Of_The_Freshwater_Zooplankton_Genus_Holopedium_Crustacea_Branchiopoda_Ctenopoda_Revealed_By_Genetic_Methods/links/5b22930b0f7e9b0e374325fd/Three-New-Cryptic-Species-Of-The-Freshwater-Zooplankton-Genus-Holopedium-Crustacea-Branchiopoda-Ctenopoda-Revealed-By-Genetic-Methods.pdf

Fotos / Sonidos

Observ.
lamawebberDescripción
Thalassiosira nordenskioeldii Cleve 1873
SEM images of the marine diatom Thalassiosira cf. nordenskioeldii (Castracane) Hasle 1983. Phylum: Bacillariophycophyta, Subphylum: Bacillariophytina, Class: Mediophyceae, Order: Thalassiosirales, Family: Thalassiosiraceae. Girdle view. Thalassiosira nordenskioeldii cells are cylindrical to discoid and usually joined together in long chains. In girdle view T. nordenskioeldii cells appear as octagonal discs. They have a pronounced concavity at the valve center from which a thick mucilage tube connects to the centre of a neighbouring cell. Mantle edges are beveled. Long radiating mucilage threads exit from the prominent marginal strutted processes at the bend between the valve face and the mantle.
Cell diameter is recorded between 10-50 µm in diameter (Horner 2002: 29).
Abundant, especially during the spring bloom in the Trincomali Channel. Collected by a 20 µm plankton net April 6, 2022 from the Spanish Hills Wharf (N48˚ 59.688’, W123˚ 35.064’), Trincomali Channel, Galiano Island, Southern Gulf Islands, British Columbia, Canada.
Imaging with a Hitachi s4800 and TM4000 SEMs at the AMF at UVIC. Thank you Ron Read for taking some of the SEM images and Elaine Humphrey for SEM support. Adjusted in PhotoShop.
References:
Cupp, E. E. 1943. Marine Plankton Diatoms of the West Coast of North America. University of California Press. Berkeley, California.
Hoppenrath, M., Elbrachter, M., Drebes, G. (2009) Marine Phytoplankton, Selected microphytoplankton species from the North Sea around Helgoland and Sylt. pp. 59, figure 5. Plate 24, page 58:fig. f-k. E. Schweizerbart’sche Verlagsbunchhandlung, Stuttgart, Germany.
Horner, R. A. (2002), A Taxonomic Guide to Some Common Marine Phytoplankton. p. 31. Biopress Ltd., Bristol.
Hasle, G.R. & Syvertsen, E.E. (1996). Marine Diatoms. In: Identifying Marine Phytoplankton. (Tomas, C.R. Eds). San Diego: Academic Press.
Guiry, M.D. & Guiry, G.M. 2007, AlgaeBase version 4.2. World-wide electronic publication, National University of Ireland, http://algaebase.org, searched April 10, 2022.
F.E. Round, F.E., Crawford, R.M. and Mann, D.G. (1990). The Diatoms, Biology & Morphology of the Genera. Cambridge University Press, Cambridge, UK. pp. 132-133.
Fryxell, G.A. and Hasle, G.R. (1979). The genus Thalassiosira: species with internal extensions of the strutted processes. Phycologia. vol. 18 (4), 378-393.
Pienitz, R., Fedje, D. and Poulin, M. (2003) Marine and Non-Marine Diatoms from the Haida Gwaii Archipelago and Surrounding Coasts, Northeastern Pacific, Canada In Bibliotheca Diatomologica (H. Lange-Bertalot and P. Kociolek, eds.), Band 48, J. Cramer, Stuttgart, 146 pp.
Shim, J. H. (1976). Distribution and Taxonomy of Planktonic Marine Diatoms in the Strait of Georgia, B.C. Phd. Thesis, UBC. Plate VII, fig. 2a & b.
Waters, R. E., Brown, L.N., and MG Robinson, M.G. (1992). Phytoplankton of Esquimalt Lagoon, British Columbia: comparison with west Vancouver Island coastal and offshore waters. Canadian Technical Report of Hydrography Ocean Sciences 137.